Masai Giraffe and Habitat Preference in Three Northern Tanzanian Conservation Areas
Abstract
The (Masai Giraffe) Giraffa c. Tippelslirichi was surveyed for habitat preference-activity relationships using structured index classes. Specific comparison of activity patterns; feeding, walking and standing among grassland and open brush land/shrub land habitat preference in Tarangire National Park and Manyara Ranch Conservancy, resulted in moderate association. The study was conducted at different total site areas and during the dry season with spatial geo-located occurrences.
Introduction
The predominant research conducted on Giraffe for the past few years to the present has focused on the species morphological, behavioral and social attributes. Of the eight main species studied, Giraffa c. Tippelslirichi is the most wide spread in Tanzania (Kingdon 1997). Most of these studies cover social unit structure and activity pattern which can depend on sex with adult females holding wide home ranges and adult males forming smaller territorial ranges. These works also touch on critical adaptive morphology of the Giraffes long neck for browsing and its long visual capacity with a strong retina. In functional terms, the horizontal retina streak improves visual sampling across the horizon, thereby allowing detection of predator, prey, or con-specifics without the need for an extensive head (Coimbra et. al, 2013).
However this species focused work is rarely studied in relation to habitat resources or spatial classification. A common research subject when exploring the topic of habitat resources and wildlife conservation management is carrying capacity. This resource management idea, that an area can support only so many animals is well established. In a management context its used to determine the highest number of animals a habitat can maintain in a healthy, vigorous condition. Carrying capacity is not a fixed factor of the land and will change as food, cover, and water change with the seasons or over long time periods (Patton 1997).
Continuous management is essential when managing wildlife carrying capacity and any of these indirect effects on species like Giraffe activity should be assessed. Today such habitat and activity exploratory/comparison research is more feasible due to geomatics technology. By using remote sensing and practical surveying, our study investigated Giraffa c. Tippelslirichi (Masai Giraffe) habitat preference and whether an associated relationship with activity in three Northern Tanzanian conservation areas exists during the dry season. For management purposes the research also discusses spatial components of habitat use.
Methodology
The Study Area
The study was conducted within three Northern Tanzanian wildlife conservation areas of Lake Manyara National Park (LMNP) (3° 29’ S; 35° 50’ E), Tarangire National Park (TNP) (3° 59’ S; 36° 00’ E) and Manyara Ranch Conservancy (MRC) (3° 35’ S; 36° 00’ E). These wildlife areas all exist in portions of Arusha-Manyara-Shinyanga Region of Tanzania which holds a regional temperature of (13–30°C) (Winks 2009). The Lake Manyara National Park contains largely dense broad-leaf forest and several seasonal/non-seasonal springs. While Tarangire National Park contains largely bushland/shrubland and Manyara Ranch Conservancy partially comprised of grassland/woodlands.
Geologically the region’s land forms are of volcanic origin, with a heterogeneous mixture of the following soils; chernozems, leptosols, nitisols, luvisols and andosols. The area has an arid to semiarid climate which varies with altitude, season, terrain and also includes sub-humid forests in hydro-logically active regions. Rainfall ranges between 400 and 900 mm years -1 (Winks 2009). The regional microbiome of Northern Tanzania as a whole is comprised of fragmented grasslands, bush lands, shrub lands, woodlands and broad leaf forests depending on abiotic factors. The regional wildlife most importantly Giraffe may inhabit these different vegetation types based on social and physical evolutionary niches to micro-habitat.
Experimental Set-Up
The total study area with the three conservation sites combined was approximately 3000 km 2. A randomized observational survey of Giraffa c. Tippelskirichi (Fig. 1) was conducted at a distance of 30–200 m using activity and vegetation type classifications. The following activity index classes were determined with the corresponding numbers for identification: feeding 1, standing 2, Vigilant 3, Walking 4, Mating 5 and likewise for the following habitat classes: woodland 1, Bushland/Shrubland Open 2, Bushland/Shrubland Closed 3 and Grassland 4 (Kioko 2016).

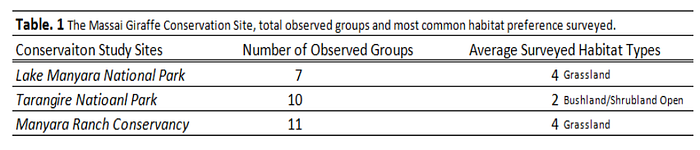
The LMNP a 9 km 2 area was surveyed on July 15, 2016 from noon to four o’clock, TNP a 2849 km 2 area on July 19, 2016 from ten to four o’clock and MRC a 142 km 2 area on July 20, 2016 from nine to two o’clock. All surveying took place in similar weather of full sun. The data from each conservation area was compared by measuring the frequency and average habitat type preferred among each surveyed site (Table. 1). In addition, data of giraffe activity and habitat use from the two sites with alike habitat preference (TNP & MRC) were compared spatially using remote sensing and GPS technology for assessing conservation area-habitat relationships.
The design was intended to be randomized to remove bias error in sampling. To accomplish this, representative samples were collected in a spontaneous route during a similar range of time during the day and all in the dry season.
Results
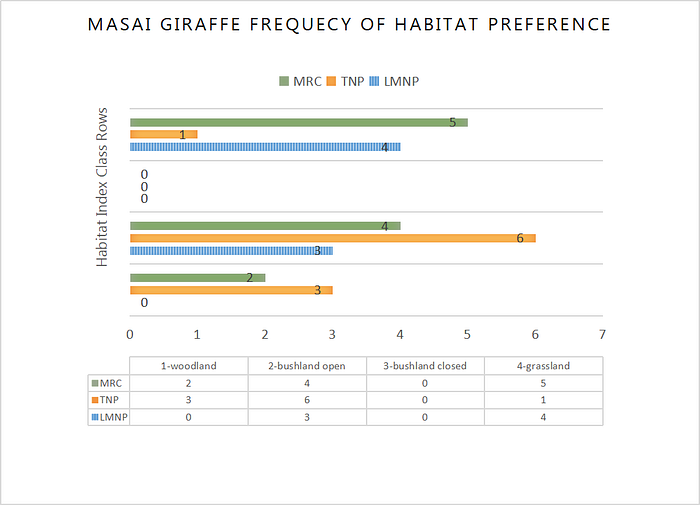
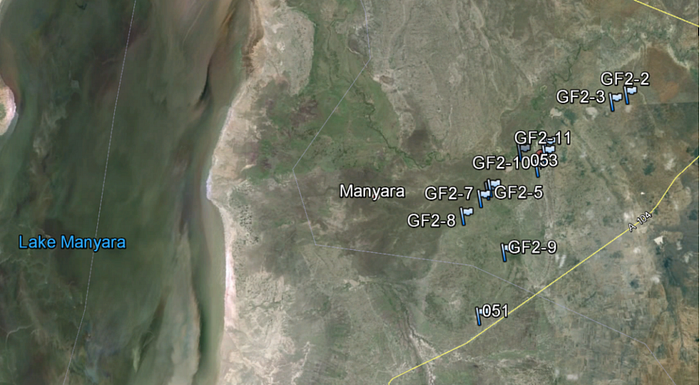
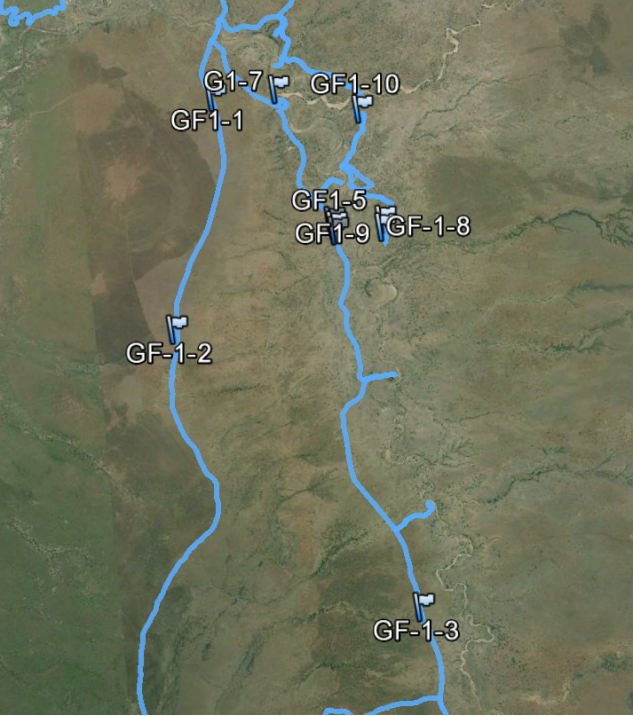
Data analysis from the three surveyed sites indicates Giraffa c. Tippelskirichi predominately engages in feeding, walking or stance activities within the three Northern Tanzanian study sites (Fig. 2). Of the four habitat index classes, the species was observed to be occupying mostly Grasslands and open Bushland/Shrubland within the three conservation areas. When observational groups among the three study sites were compared with habitat use, a similarity occurred with both high average use of 4-grasslands and frequency of 2-open bushland habitat preference among Tarangire National Park, Manyara Ranch Conservancy and Lake Manyara National Park (Table.1). The highest number of observed groups were surveyed in Manyara Ranch Conservancy (MRC) and lowest in Lake Manyara National Park (LMNP). With most of LMNP observed by remote sensing to be broad-leaf forest and MRC being mostly grassland, its understandable that of the two conservation area ecosystems MRC provides more of a suitable habitat for feeding, walking and standing in safety (appendix. 1). The Giraffe densities in Manyara are high compared to other areas within the Masai ecosystem, and Manyara probably serves as a dry season refuge (Jeugd 2000).
Discussion
To understand the background behind habitat preference and activity pattern relationships, its important to consider factors of feeding ecology. The Giraffe is an ideal animal for examining questions of feeding ecology. First because its long neck allows it to reach foliage unavailable to most other herbivores (Truman et. al, 1991). This ecological feeding niche provide the Giraffe with access to food at a strata of 5 meters without any substantial feeding competition by other species.
A sex ratio pattern among feeding habitat areas for Giraffe is also apparent. Males are more common in forest and in taller, thicker vegetation and females are more common in open habitat (Truman et. al, 1991). This sex ratio phenomenon was present among the study areas, with a higher portion of female groups surveyed in grassland habitats and fewer male coalitions or individuals observed. Its been suggested that some reasons for this different feeding strategy in genders, is to remove inter-specific competition for food and that most males are 20% taller than females providing a greater field of vision and overall security within foliage dense habitats. With mean of home range area estimates of 282 km 2 for Giraffe (du Toit 1990).
Spatial scaling in mammals has also been investigated comparing home range area with metabolic rate. Hence, larger animals appear to feed over a proportionally greater area than is predicted from metabolic requirements alone. The changing season and available plant composition effects this feeding and metabolic requirement in a major way. Over 100 different plants have been recorded in the giraffes diet, but various species of acacias and Combretum form the bulk of their diet in most areas. Giraffe are exclusive browsers, mainly focusing on nutritious young and new tree and shrub shoots, but Acacia trees are their favorite choice of forage (Okello et. al, 2015).
Like many savanna herbivores, they disperse widely during the rains and concentrate along watercourses in the dry season, were high quality forage continues to be available (Estes 1991). These Masai Giraffe feeding and habitat use patterns near water sources are present spatially in the Lake Manyara National Park (appendix. 1). In addition grasslands and open bushlands/shrublands of Northern Tanzania where most Masai Giraffe species were observed, is dominated by acacia stands. So based on these observed habitat preferences of Giraffe and their related feeding, walking and standing activities, its concluded that Tarangire National Park and Manyara Ranch Conservancy hold a moderate species habitat to activity relationship and Lake Manyara National Park a much weaker relationship. This comparative information can prove to be vital when preparing and implementing conservation management strategies for Giraffa c. Tippelskirichi in a stand alone wildlife area or plans for multiple conservation areas.
Conclusion
This survey study during the dry season in Northern Tanzania presented a noticeable relationship in Giraffe habitat preference and activity pattern among Tarangire National Park and Manyara Ranch Conservancy.
The distinct feeding ecology and sex ratio characteristics in habitat use also support spatial patterns observed using remote sensing.
When managing the Giraffa c. Tippelskirichi in Northern Tanzania conservation areas its suggested these study results be accounted for in implementation and preparation.
Its also recommended that data collection be continued for this study and that Giraffe to plant specific interactions be analyzed further.
Acknowledgments
This study would not be possible without The School For Field Studies research directors and staff. The conservation area community and rangers in the respective parks also contributed immensely to the implementation of this study.
Appendix
References
Blomley, T., Peliegner, K., Isango, J., Zahabu, E., Ahrends, A., & Burgess, N. (2008). Seeing the wood for the trees: An assessment of the impact of participaroty forest management on forest condition in Tanzania. Oryx. Oryx, 42(3), 380–391.
Briggs, P. (1993). Guide to Tanzania. Old Saybrook, Connecticut: Bradt Publications.
Carter, K. D., Frere, C. H., Seddon, J. M., Carter, J. K., & Goldizen, A. W. (2013). Fission-fusion dynamics in wild giraffes may be driven by kinship, spatial overlap and individual social preference. Animal Behaviour, 85, 385–394.
Coimbra, J. P., Hart, N. S., Collin, S. P., & Manger, P. R. (2013). Scene From Above: Retinal Ganglion Cell Topography and Spatial Resolving Power in the Giraffe. The Journal of Comparative Neurology, 521, 2042–2057.
Estes, R. D. (1991). African Mammals. Los Angeles, California: University of California Press Berkley.
Jeugd, H. V. (2000). Movement and group structure of giraffe (Giraffe camelopardalis) in Lake Manyara National Park, Tanzania. Journal of Zoology, 251, 15–21.
Kingdon, J. (1997). The Kingdon Field Guide To African Mammals. Wiltshire, United Kingdom: Princeton University Press.
Meissner, C. H., & Schultheiss, W. A. (1995). The nutritive value of veld as indicated by faecal phosphorous and nitrogen and its relation to the condition and movement of prominent ruminants during the 1992–1993 drought in the Kruger National Park. Koedoe, 38(1), 17–31.
Okello, M. M., Kenana, L., Maliti, H., Kiringe, J. W., Kanga, E., Warinwa, F., . . . Mwita, M. (2015). Population Status and Trend of the Maasai Giraffe in the Mid Kenya-Tanzania Borderland. Natural Resources, 6, 159–173.
Omphile, U. J., & Powell, J. (2002, July). Large ungulate habitat preference in Chobe National Park, Botswana. Journal of Range Management, 55, 341–349.
Patton, D. R. (1997). Wildlife Habitat Relationship In Forested Ecosystems. Portland, Oregon: Timber Press.
Pellew, R. A. (1984, April). Food Consumption and Energy Budgets of the Giraffe. The Journal of Applied Ecology, 21(1), 141–159.
Prins, H. H., & Der Jeugd, H. P. (1993). Herbivore populaiton crashes and woodland structure in East Africa. Journal of Ecology, 81, 305–314.
Thomassen, H. A., Freedman, A. H., Brown, D. M., & Buermann, W. (2013, October). Regional Differences in Seasonal Timing of Rainfall Discriminate between Genetically Distinct East African Giraffe Taxa. PLOS ONE, 8(10), 1–14.
Toit, J. D. (1989). Body size, population metabolism, and habitat specialization among large African herbivores. American Naturalist, 133, 736–740.
Toit, J. D. (1990). Home range — body mass relation: A field study on African browsing ruminants. Oecologia, 85, 301–303.
Truman, Y. P., & Isbell, L. A. (1991). Sex Differences in Giraffe Feeding Ecology: Energetic and Social Constraints. Ethology, 87, 79–89.
Valeix, M., Loveridge, A. J., Chamaille-Jammes, S., Davidson, Z., Murindaggomo, F., & Fritz, H. (2009). Behavioral adjustments of African herbivores to predation risk by lions: Spatiotemporal variations influence habitat use. Ecology, 90(1), 23–30.
Waltert, M., Meyer, B., & Kiffner, C. (2009). Habitat availability, hunting or poaching: What affects distribution and density of large mammals in western Tanzanian woodlands. African Journal of Ecology, 47, 737–746.
Winks, Q. (2009). The essential guide to customs & culture Tanzania. New York, New York: Culture Smart.
